

ICU Physiology in 1000 Words: Pulmonary Vascular Resistance & Linear Assumptions

Jon-Emile S. Kenny MD [@heart_lung]
A recent review of Paul Wood’s consummate approach to pulmonary hypertension is well-worth a read [1]. In it, the basic physiology of the calculated pulmonary vascular resistance [cPVR] is presented, as it often is, as a linear function. Recall, maybe traumatically, the grade-school equation y = mx + b. With this ontology, we presume that x and y are the independent and dependent variables, respectively. In other words, x is systematically varied while y is ‘spit out’ of the mathematical machine after modified by the slope ‘m’ and the y-intercept ‘b’.
How does this concern the cPVR? As the relationship is presented, mean pulmonary arterial pressure [mPAP] equals the cPVR times cardiac output [Q] plus the left atrial pressure [taken to equal the pulmonary artery occlusion pressure, Ppao]. That is, mPAP = cPVR x Q + Ppao [i.e. y = mx + b]. Put another way, the mPAP results by adding to the Ppao the product of cardiac output and the slope, cPVR [Figure 1].
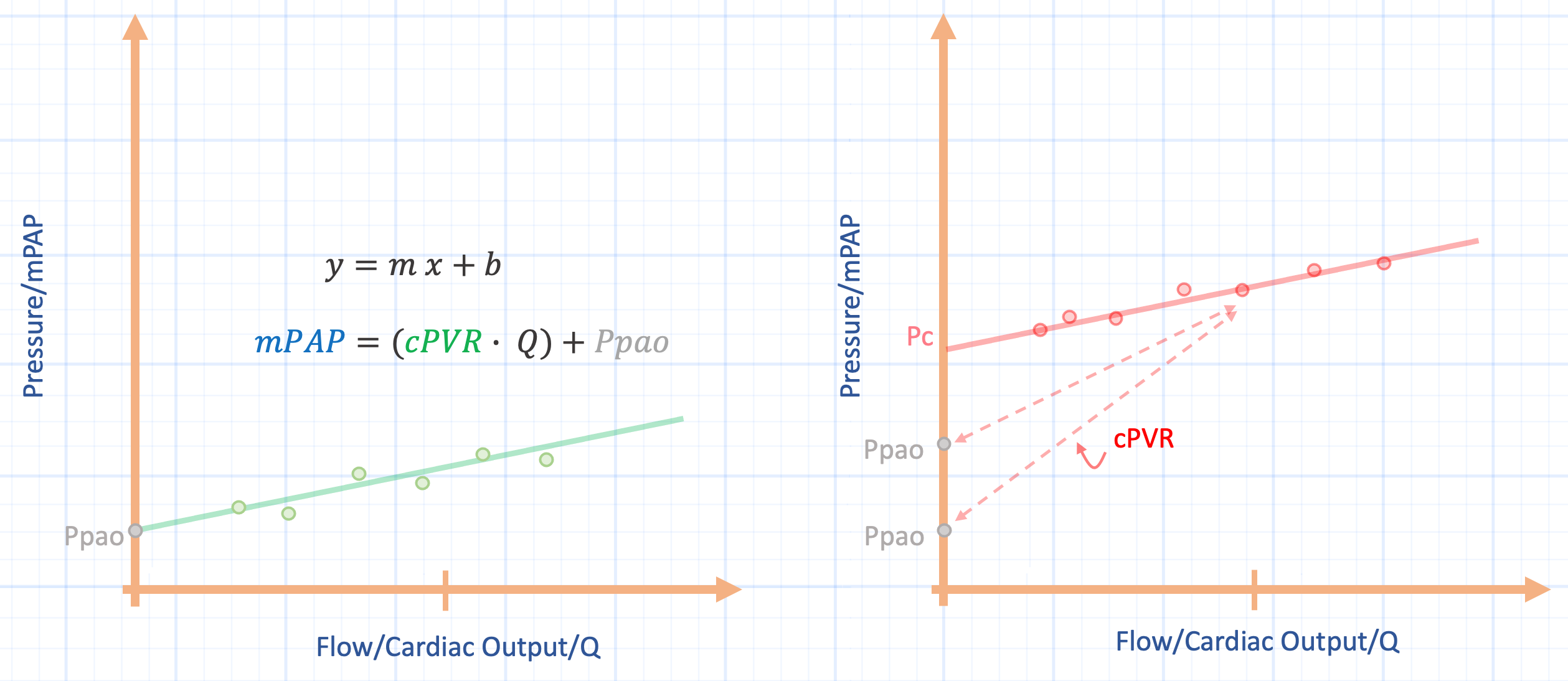
Figure 1: Pulmonary vascular pressure-flow plots. x-axis is flow, or Q and y-axis is pressure, or mean pulmonary arterial pressure [mPAP]. Left panel shows the linear approximation for a series of pressure-flow points at a constant pulmonary artery occlusion pressure [Ppao]. Right panel shows how the calculated pulmonary vascular resistance is over-estimated when the closing pressure [Pc] is much greater than the Ppao. The Ppao approaches the Pc with left atrial hypertension, for example, this causes the cPVR to fall [diminishing slope].
The slope of the relationship, cPVR, is often thought to represent active vascular constriction and dilation – set physiologically by vascular ‘tone’ – and that measuring mPAP, Q and Ppao simply reveal to us this underlying truth of the cPVR. Might changing Q or Ppao, by themselves, alter the slope of the relationship without any change in vascular tone? Do other biophysical properties mediate the cPVR and are there measurement assumptions to consider?
‘Recruitment’
Inherent in the aforementioned model is that the Ppao represents the left atrial pressure and that these values truly act as the ‘downstream’ pressure for pulmonary blood flow. This is true when the vasculature comports with ‘West zone’ III – when the transmural vascular distending pressure is larger than any opposing pressure from the pulmonary artery through to the left atrium [2]. Yet, when the lung – or portions of it – lie within zone I or II conditions, the left atrial pressure is not the downstream pressure, something called the ‘closing pressure’ [Pc] becomes the effective pressure sink [3-6]. In other words, the pressure gradient is not mPAP less Ppao, but rather the mPAP less the Pc. This type of physiology typically occurs in the upright, gravity dependent lung, but can also occur in common disease states such as pulmonary embolism, acute respiratory distress syndrome [7] or excessive airway pressure [8, 9].
When the Ppao is significantly lower than the Pc, the cPVR overestimates ‘vascular tone’ for the lung [Figure 1, right panel]. It is challenging to obtain the Pc in the human lung as it requires measuring multiple mPAP-Q coordinates at a constant left atrial pressure followed by extrapolating the linear relationship to the pressure axis [6]. Typically, the Pc is greater than the Ppao; in the canine lung, the Ppao does not approach the Pc until roughly 19 mmHg [3]. This physiology may explain falling cPVR in both human and animal studies where left atrial pressure acutely rises [10, 11]. In some ways this is akin to pulmonary vascular ‘recruitment’ in that vessels in which the Pc is greater than the Ppao are opened by rising downstream pressure; the cPVR falls, but this does not necessarily mean that vessels have ‘dilated’ by vascular smooth muscle relaxation [3].
‘Dilation’
The abovementioned representation of the pulmonary vascular tree is sometimes termed the ‘Starling’ or ‘waterfall’ model [12, 13]. In addition to being ‘recruited’ [e.g. by opening a Starling resistor], pulmonary blood vessels can also passively ‘dilate’. Passive vascular dilation also alters the slope of the mPAP-Q relationship, that is, change the cPVR. More simply, even if the Ppao and Pc are assumed to be equal and constant, increasing blood flow flattens the slope of the cPVR because vessels dilate as their volume rise; thus, blood flow and cPVR are functionally linked. Both ‘recruitment’ and ‘dilation’ as described are passive processes and explain why the mPAP-Q relationship is not entirely linear, especially at low and high levels of Q [14].
Mathematical models of cPVR that include inherent, passive vascular distensibility have been proposed and better fit experimental data [13]. For example, at high blood flow, linear extrapolations to estimate Pc are inaccurately elevated [6]; equations that factor in distensibility better pinpoint pulmonary vascular closing pressure. Notably, the distensibility constant [Figure 2, alpha] employed in these equations is quite similar across species. Within humans, there is some minor differences between men and women and with aging [6].
‘Stiffening’
While the pulmonary vasculature can ‘recruit’ and ‘dilate’ as above, can these passive processes reach a tensile limit? Interestingly, the volume-pressure relationship of pulmonary arteries and veins is slightly different [15, 16]. Most pulmonary veins, and small arteries, reach a plateau around 19 mmHg – where further increase in volume results in proportionally more pressure elevation [17] [Figure 2, inset]. As previously argued, this may be why cPVR rises as a function of vessel ‘stiffening’ rather than vascular constriction. Indeed, in a canine model of chronic pulmonary venous hypertension, cPVR rose without an increase in the pulmonary arterial diastolic – Ppao pressure gradient [10]. Further, in this model vascular elastance [stiffness] and wave velocity both increased. These canine data may explain why roughly one-third of patients with isolated ‘post-capillary’ pulmonary hypertension had a cPVR above 3.0 [18] and why changes in Ppao are directly related to pulmonary arterial input impedance and indirectly related to pulmonary vascular compliance in humans [11] [Figure 2].

Figure 2: When distensibility [alpha] is factored into the pulmonary vascular resistance. 2% alpha means that for each 1 mmHg in pressure, vascular diameter rises by 2%. Note for a constant flow [x-axis], decreasing distensibility [0% alpha] increases the cPVR for constant Ppao. The right inset shows that as vascular pressure rises, vascular volume begins to plateau, that is, distensibility falls. This, very generally, occurs around 20 mmHg.
Conclusions
Passive pulmonary vascular recruitment and dilation can lower the cPVR independent of vasomotor tone. Thus, falling cPVR does not necessarily imply smooth muscle dilation, but could simply reflect rising Ppao and/or flow. Further, and especially in chronic venous pulmonary hypertension, cPVR may elevate when pulmonary vessels are dilated to their tensile limits. Caution is advised when inferring active changes in the pulmonary circulation from only cPVR; linear approximation is imperfect and subject to other biophysical forces.
Please check out other posts in this series,
JE
Dr. Kenny is the cofounder and Chief Medical Officer of Flosonics Medical; he also the creator and author of a free hemodynamic curriculum at heart-lung.org. Download his free textbook here.
References
Newman JH: Pulmonary Hypertension by the Method of Paul Wood. Chest 2020, volume 158, issue 3, P1164-1171.
West JB, Dollery CT, Naimark A: Distribution of Blood Flow in Isolated Lung; Relation to Vascular and Alveolar Pressures. Journal of applied physiology 1964, 19:713-724.
Ducas J, Schick U, Girling L et al: Effects of altered left atrial pressure on pulmonary vascular pressure-flow relationships. American Journal of Physiology-Heart and Circulatory Physiology 1988, 255(1):H19-H25.
Duval D, Ducas J, Molloy W et al: Effects of pulmonary (P) emboli (E) and noradrenaline (NE) on pulmonary pressure-flow relationships. Am Rev Respir Dis 1984, 129(4):A63.
Ducas J, Duval D, Dasilva H et al: Treatment of canine pulmonary hypertension: effects of norepinephrine and isoproterenol on pulmonary vascular pressure-flow characteristics. Circulation 1987, 75(1):235-242.
Naeije R, Vachiery JL, Yerly P et al: The transpulmonary pressure gradient for the diagnosis of pulmonary vascular disease. The European respiratory journal 2013, 41(1):217-223.
Leeman M, Lejeune P, Closset J et al: Nature of pulmonary hypertension in canine oleic acid pulmonary edema. Journal of Applied Physiology 1990, 69(1):293-298.
Lopez-Muniz R, Stephens N, Bromberger-Barnea B et al: Critical closure of pulmonary vessels analyzed in terms of Starling resistor model. Journal of Applied Physiology 1968, 24(5):625-635.
Zhuang FY, Fung YC, Yen RT: Analysis of blood flow in cat's lung with detailed anatomical and elasticity data. Journal of applied physiology: respiratory, environmental and exercise physiology 1983, 55(4):1341-1348.
Hopkins RA, Hammon Jr JW, McHale PA et al: An analysis of the pulsatile hemodynamic responses of the pulmonary circulation to acute and chronic pulmonary venous hypertension in the awake dog. Circulation research 1980, 47(6):902-910.
Tedford RJ: Determinants of right ventricular afterload (2013 Grover Conference series). Pulmonary circulation 2014, 4(2):211-219.
Mitzner WA: Resistance of the pulmonary circulation. Clinics in chest medicine 1983, 4(2):127-137.
Linehan JH, Haworth ST, Nelin LD et al: A simple distensible vessel model for interpreting pulmonary vascular pressure-flow curves. Journal of Applied Physiology 1992, 73(3):987-994.
Sylvester JT, Mitzner W, Ngeow Y et al: Hypoxic constriction of alveolar and extra-alveolar vessels in isolated pig lungs. Journal of applied physiology: respiratory, environmental and exercise physiology 1983, 54(6):1660-1666.
al-Tinawi A, Madden JA, Dawson CA et al: Distensibility of small arteries of the dog lung. Journal of Applied Physiology 1991, 71(5):1714-1722.
al-Tinawi A, Clough AV, Harder DR et al: Distensibility of small veins of the dog lung. Journal of Applied Physiology 1992, 73(5):2158-2165.
Maloney JE, Rooholamini SA, Wexler L: Pressure-diameter relations of small blood vessels in isolated dog lung. Microvascular research 1970, 2(1):1-12.
Assad TR, Brittain EL, Wells QS et al: Hemodynamic evidence of vascular remodeling in combined post-and precapillary pulmonary hypertension. Pulmonary circulation 2016, 6(3):313-321.